An in-depth look at the pelvic reconstruction of Ardipithecus
The pelvic anatomy of Ardipithecus ramidus may give clues about the posture and locomotion of this ancient species.

One of the most severely crushed parts of the Ardipithecus ramidus ARA-VP-6/500 skeleton is the pelvis. The left os coxa is nearly complete but badly distorted, part of the right ilium is preserved along with a bit of the sacrum. The research team had to correct for the distortion and breakage of the os coxa to interpret its form. The resulting paper, by Lovejoy and colleagues (2009c), describes the anatomy as present in the distorted fossil, the CT-assisted reconstruction of the fossil, and a whole-pelvis model based on the reconstruction.
I've written some thoughts on the specimen and its reconstruction. These are not nearly comprehensive, but enough to introduce a bit of the complexity of the locomotor inference from this odd pelvis. Also, I've pointed to the one Miocene ape notably absent from the published comparisons, Oreopithecus.
The pelvic model
I want to spend some time considering their 3-d pelvic model. I find it to be a provocative application of visualization technology. On the one hand, a model greatly facilitates the description and comparison of the specimen with other key fossils. But on the other hand, any model comes with a load of assumptions, which may not be made explicit when anatomical comparisons are made. It’s our job to approach models skeptically, to try to identify the implicit assumptions and bring them out to be examined.
In this case, much thought has gone into the virtual model. For astute Ardipithecus-watchers, a paper published ten years ago has played a central role in understanding the probable anatomy of the specimen. Here’s the first figure from that paper, published in 1999 by Lovejoy, Martin Cohn and Tim White:
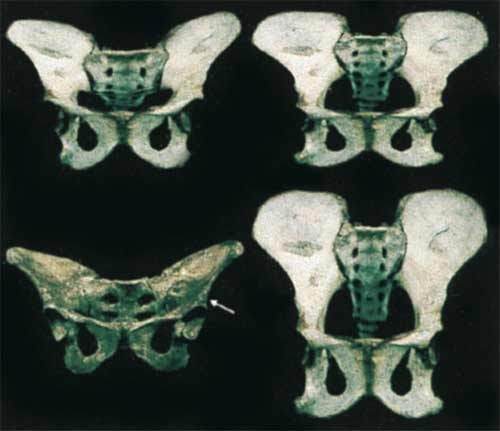
Very interesting indeed. The top left image is indeed very close to the Ardipithecus model that Lovejoy and colleagues present in the current paper. The figure (Figure 1 in Lovejoy et al. 1999) had the following, very long caption:
Fig. 1. Hypothetical transitional emergence of the hominid pelvis. Anterior photographs of a chimpanzee pelvis (lower right) and that of A.L. 288-1 (“Lucy”) (lower left) were scanned. Using a sliding scale, the upper left image was then obtained by digital morphing to a transitional stage 75% of the distance between the chimpanzee and A.L. 288-1. The upper right image is a simple superoinferior digital distortion of the chimpanzee without any reference to a known “end product” (Photoshop “scale function”). Image breadth was not altered; its superoinferior height was simply reduced by 2/3. (Note: the upper right image appears somewhat less “transitional” than the one at the upper left because the latter benets from the three-dimensionality of the two images being morphed; i.e., our distortion was only two-dimensional.) We do not suggest that either image constitutes an actual “intermediate” pelvic form. We wish only to demonstrate that a simple dimensional change in one hypothetical adult form is very similar to that which has been morphed by using the known adult “final outcome” and that it might be achieved by a simple underlying mechanism such as a progressive increase or decrease in the slopes of cell response gradients (see text). We suggest that this is the most probable morphogenetic mode by which the many anatomical differences between A.L.-288-1 and the chimpanzee pelves evolved. Therefore, the isolated denition and separate analysis of each of the many traits that differ between these pelves is likely to greatly distort their functional and phyletic signicance (see especially ref. 7, pp. 359–361 for discussion). Note, for example, that a number of the unusual distinguishing characters of the australopithecine pelvis, including its exceptionally broad sacrum, platypelloid birth canal (i.e., anteroposterior dimension/mediolateral dimension × 100 ≈ 50–60), short pubic symphysis, elongated superior and inferior pubic rami, ovoid obturator foramina, etc., have all been reproduced by this simple, relatively crude, linear distortion (Lovejoy et al., 1999, 13249).
OK, so the top right image is a squashed chimpanzee pelvis; the top left is the product of “morphing” the chimpanzee pelvis with Lucy’s pelvis. Lovejoy et al. (1999) were making a simple point: you can get almost all the features of the hominid pelvis by a very simple process of morphological change. The model of the pelvis was an illustration for a developmental model—the idea was that many morphological features of the hominins might result from a few changes to gene regulation early in development.
In the current paper, the model is no longer a mere thought experiment. In 1999, readers of the paper could hypothesize that Lovejoy and colleagues already knew the morphology of the Ardipithecus pelvis—Tim White’s famous “Star Wars” comment was a clue. But in the current paper (Lovejoy et al., 2009c), it seems clear that the 3-d digital version of the model also shaped the reconstruction of the highly crushed pelvic remains. That means we have to carefully read Lovejoy and colleagues’ (2009) paper to determine which parts of the model are fully supported by the specimens’ preserved (non-crushed) anatomy, and which are more speculative.
The ilia
One aspect of the digital model got my attention more than the others, because it was not evident in the 1999 pelvis model. It’s not the length or breadth of the ilium—based on the preserved bone, they have reconstructed both as intermediate between the chimpanzee and australopithecine morphology. Nor is it the chimpanzee-like long ischium, without which it’s hard to see how Ardipithecus could have taken much advantage of those grasping feet. No, to me the surprising thing is that they’ve given the iliac blade a pronounced anterior cant, far more so than in Australopithecus.
The best view of this aspect of the reconstruction is the supplementary figure S1 from Lovejoy and colleagues 2009c, which presents the Ardi reconstruction superimposed in space on the reconstructed pelvis of Lucy.
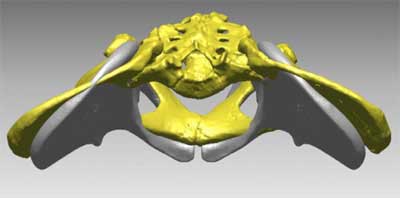
You can see that the grey Ardi pelvis has ilia that angle far more anteriorly than Lucy’s. This is the opposite direction from any of the living great apes, which have iliac blades aligned more parallel to the back of the trunk.
The description does not provide enough evidence to make it clear to what extent this position is necessary based on the preserved morphology. The os coxae are crushed, and the paper does not illustrate the parts in sufficient detail to for me to judge the reconstruction. It appears to me that the position is determined by two assumptions. First, they provide the specimen with a wide sacrum. In the comparison between the Ardi model and Lucy’s pelvic reconstruction, it is clear that they’ve fitted Ardi with the same proportionate size sacrum as Lucy, which—since the comparison is with a Lucy blown up to 115% life-size—is 15 percent wider than Lucy’s. Second, they key the position of the anterior superior iliac spine off that of the anterior inferior iliac spine. Since this anterior inferior spine is especially prominent on Ardi’s os coxa, keeping the two spines in their usual conformation would require an anteriorly placed anterior superior iliac spine.
Of course, the only extant hominoids to assess the relation between these spines are highly suspensory apes with stiff, short lumbar spines, and us: short-pelvised obligate bipeds. If Ardipithecus was neither of these—and it wasn’t—then maybe it had an unexpected relation between these anatomical features.
At any rate, the shape reconstructed for Ardipithecus goes along with Lovejoy and colleagues hypothesis about ilium and lumbar spine evolution. They propose that the ilia have flattened and extended in length independently in at least the three lineages of chimpanzees, gorillas, and orangutans (and we may add gibbons) as a consequence of sacralizing the lower lumbar vertebrae and reducing the flexibility of the lower back, thereby creating a short, stiff trunk.
Given that scenario, we can’t consider the ilia apart from the lumbar spine and some assumptions about the requirements of different locomotor patterns. Unfortunately, Ardipithecus doesn’t have any lumbar vertebrae — or indeed any sacrum worth reconstructing.
But there is another Miocene ape that does.
The relevance of Oreopithecus
The best-preserved pelvic remains from any Miocene ape are those of Oreopithecus bambolii. This species has been found in lignite deposits of Tuscany and Sardinia, dating to between 10 and 7 million years ago. At that time, these regions were joined as a single landmass, an island not connected to the European mainland, making it credible (though not certain) that Oreopithecus faced lower predation than mainland primates.
I’m not going to do any kind of detailed description here of the Oreopithecus pelvic remains. A review of the Oreopithecus phylogenetic status by Terry Harrison (1986) gives a literature review to that point, more recent papers by Rook and colleagues (1999) and Kohler and Moyà-Solà (1997) took up the problem of possible habitual bipedality.
It has been evident for a long time that several features of the Oreopithecus pelvis are shared with hominins and not with living great apes. Harrison (1991) gives a list of some of these features, focusing on the importance of the anterior inferior iliac spine:
The ilium of the pelvis is quite short and broad by comparison with most extant primates, and the iliac blades are laterally flaring and coronally aligned. The anterior inferior iliac spine is prominent. Below the spine, and located just above the superior rim of the acetabulum, is a large, roughened triangular depression. Together these provide the sites of attachment for the head(s) of m. rectus femoris muscle and the ilio-femoral ligament, which were apparently well-developed. As noted by several previous workers, the broad ilium and the prominent anterior inferior spine are features by which Oreopithecus more closely approaches the hominid condition than that seen in extant great apes (Straus, 1963; Hrzeler, 1968). These specializations are functionally associated with improving the stability at the hip joint when fully extended, and for increasing the mechanical advantage of the thigh flexors and knee extensors in upright postures (Harrison, 1991, 240).
We can add to Harrison’s list other features recognized by earlier and later workers, including a short symphysis, a relatively short sacral articulation, increased breadth of the lateral ilium relative to the medial (sacral) portion, and a network of cancellous bone similar in some ways to humans.
Some argue that the biped-like anatomy of Oreopithecus is there because Oreopithecus was, in fact, a terrestrial biped. The pelvic features combine with others across the skeleton, including a lordotic lumbar spine and an extremely adducted hallux, which is proposed to give a tripod-like stability to the foot in bipedal stance (Kohler and Moyà-Solà, 1997).
I don’t necessarily share that conclusion, nor do I think that Oreopithecus was the ancestor of hominins. It evolved on an island in the middle of the ancient Mediterranean, and apparently became extinct once a land bridge connected the island with Europe. Other aspects of the skeleton, including the skull and limb proportions, show that Oreopithecus was very different from Ardipithecus.
Still, two Late Miocene hominoids with short, broad ilia and prominent anterior inferior iliac spines, is one too many. Here's a picture of the most complete Oreopithecus pelvis:
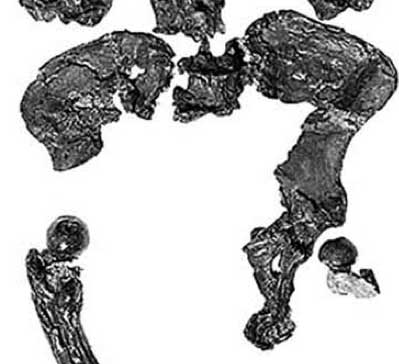
And here's the unreconstructed Ardipithecus os coxa in medial view:
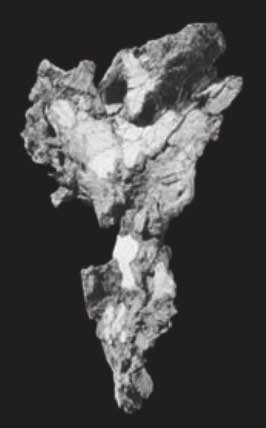
We can hypothesize a generalized ape ancestor, from which the specializations of today’s great apes and those of bipeds might arise with high likelihood under alternative selection regimes. This idea is far from new. Before the early 1960’s, the human-chimpanzee clade was a minority viewpoint, and many scientists were looking for quadrupeds that might show that some of the “bipedal” features of our pelves were primitive instead of derived.
Here’s John Robinson, writing in 1964:
In respect of the preadaptive phase [viz., before hominins became habitual bipeds], it is of great significance that Oreopithecus had a somewhat shortened innominate with a relatively broad ilium (Schultz 1960). Not only is the ilium broad, but the increased breadth is posteriad, in the region of the sacroiliac articulation. Precisely the required changes required [sic] to bring about the changed function of gluteus maximus were thereore in progress in Oreopithecus and the early Pliocene horizon at which it occurred (Hrzeler 1958) would place it at about the right period in time. From the point of view of the pelvis, therefore, Oreopithecus is remarkably suitable as an Australopithecine ancestor in the stage before the adaptive shift to erect posture occurred (Robinson, 1964, 403).
For Robinson, Oreopithecus was no more bipedal than many living apes, but lacked their extreme specializations. This made it a possible representative of the ancestral condition, somewhat more hominin-like than any of the living great apes.
I am inclined to think that’s exactly what Ardipithecus is showing. The similarity of Oreopithecus, Ardipithecus, and ultimately Australopithecus in these pelvic features suggests that this pelvic anatomy may have been the generalized form, with chimpanzees, gorillas, and orangutans separately derived toward a longer and flatter pelvis. These features would have given the large suspensory apes more stability in the lumbar region, short in living great apes, but long in both Oreopithecus and hominins.
The small acetabulum
Lovejoy and colleagues (2009c) report upper and lower bounds on acetabulum height for Ardi’s pelvis. These bounds more or less bracket those for Lucy’s pelvis, the upper bound for Ardi is the same value as the acetabulum height of Sts 14.
Both these australopithecine pelves belonged to individuals on the order of 30–35 kg in mass. White and colleagues (White et al., 2009) report an estimated mass for Ardi of 51 kg. Ardi’s acetabulum was at most the size of an average-sized chimpanzee’s despite the fact her mass as larger than most male chimps, and as much as 75 percent larger than female australopithecines. The low bound on acetabulum height is essentially the same as for the Proconsul os coxa, KNM-MW 13142 D.
If we’re looking for evidence of significant habitual weight transfer through the hindlimb, it just isn’t there.
I wouldn’t make overly much out of that, especially without a better estimate of acetabulum area. But it is one aspect of pelvic anatomy that we can observe that is more or less independent of the reconstruction of the ilia. It points away from the hypothesis that Ardipithecus was habitually bipedal.
Looking for a locomotor reconstruction
Notwithstanding any lingering uncertainty about the pelvic anatomy, I pretty much accept the arguments of Lovejoy and colleagues (Lovejoy et al., 2009a,b): Ardipithecus was a generalized arboreal quadruped. With the pelvis, the lingering questions are how much the anatomy suggests upright bipedality or carrying (Lovejoy’s “provisioning” model). My initial reaction was that Ardi may have used an arm-assisted bipedality in the trees. But having played with that idea a little, I just don’t see much support for it in the skeletal remains.
To test the importance of facultative bipedality in Ardipithecus, I think we’ll have to have a better model for how quadrupedality functions in an ape pelvis that is nonetheless very different from those of living great apes, including orangutans. The reconstructed limb proportions are within the range of Old World monkeys, suggesting that Ardipithecus would have been capable of palmigrade quadrupedalism. Lovejoy and colleagues (2009a) argue that the proximal ulna morphology is consistent with this form of locomotion also, like earlier Miocene apes and monkeys. Ardi’s long apelike ischia retain the long hamstrings lever arm necessary for powerful hindlimb extension in a flexed, quadrupedal position. And although apes do not have anteriorly-flaring ilia, monkeys do.
This is not to say that the locomotor pattern was entirely monkey-like, but merely that the ancestral, generalized quadrupedal ancestor did not have great ape locomotor specializations, including rapid vertical climbing and long-armed quadrumanous movement (Lovejoy et al., 2009a). These adaptations mainly make sense in a canopy forest habitat, where there are tall trunks devoid of lower-story branches and a continuous canopy. The grassy woodland habitat occupied by Ardipithecus would not have selected for the below-branch specializations of the forelimb and hand, and thereby would have given no reason for knuckle-walking as a terrestrial locomotor strategy.
References
Abitbol MM. 1995. Reconstruction of the Sts 14 (Australopithecus africanus) pelvis. Am J Phys Anthropol 96:143–158.
Harrison T. 1986. A reassessment of the phylogenetic relationships of Oreopithecus bambolii. J Hum Evol 15:541–584.
Harrison T. 1991. The implications of Oreopithecus bambolii for the origins of bipedalism. In: Coppens Y, Senut B, editors, Origine(s) de la bipédie chez les hominids, Cahiers de Paloanthropologie. Paris: Editions du CNRS. p 235–244.
Kohler M, Moyà-Solà S. 1997. Ape-like or hominid-like? the positional behavior of Oreopithecus bambolii reconsidered. Proc Natl Acad Sci USA 94:11,747–11,750.
Lovejoy CO, Cohn MJ, White TD. 1999. Morphological analysis of the mammalian postcranium: A developmental perspective. Proc Natl Acad Sci USA 96:13,247–13,252.
Lovejoy CO, Simpson SW, White TD, Asfaw B, Suwa G. 2009a. Careful climbing in the Miocene: The forelimbs of Ardipithecus ramidus and humans are primitive. Science 326:70e1–70e7.
Lovejoy CO, Suwa G, Simpson SW, Matternes JH, White TD. 2009b. The great divides: Ardipithecus ramidus reveals the postcrania of our last common ancestors with African apes. Science 326:100–106.
Lovejoy CO, Suwa G, Spurlock L, Asfaw B, White TD. 2009c. The pelvis and femur of Ardipithecus ramidus: The emergence of upright walking. Science 326.
Robinson JT. 1964. Adaptive radiation in the australopithecines and the origin of man. In: Howell FC, Bourlire F, editors, African ecology and human evolution. London: Methuen and Company, Limited. p 385–416.
Rook L, Bondioli L, Khler M, Moy-Sol S, Macchiarelli R. 1999. Oreopithecus was a bipedal ape after all: Evidence from the iliac cancellous architecture. Proc Natl Acad Sci U S A 96:8795–8799.
White TD, Asfaw B, Beyene Y, Haile-Selassie Y, Lovejoy CO, Suwa G, WoldeGabriel G. 2009. Ardipithecus ramidus and the paleobiology of early hominids. Science 326:75–86.

John Hawks Newsletter
Join the newsletter to receive the latest updates in your inbox.


,_4,4_milioni_di_anni_fa_02.jpg){kind=link}
{kind=link}