Blog articles
Four amazing Stone Age sites with wooden artifacts
From Africa, Asia, and Europe, these sites give us a rare window into the ways that organic technology shaped ancestral lives.

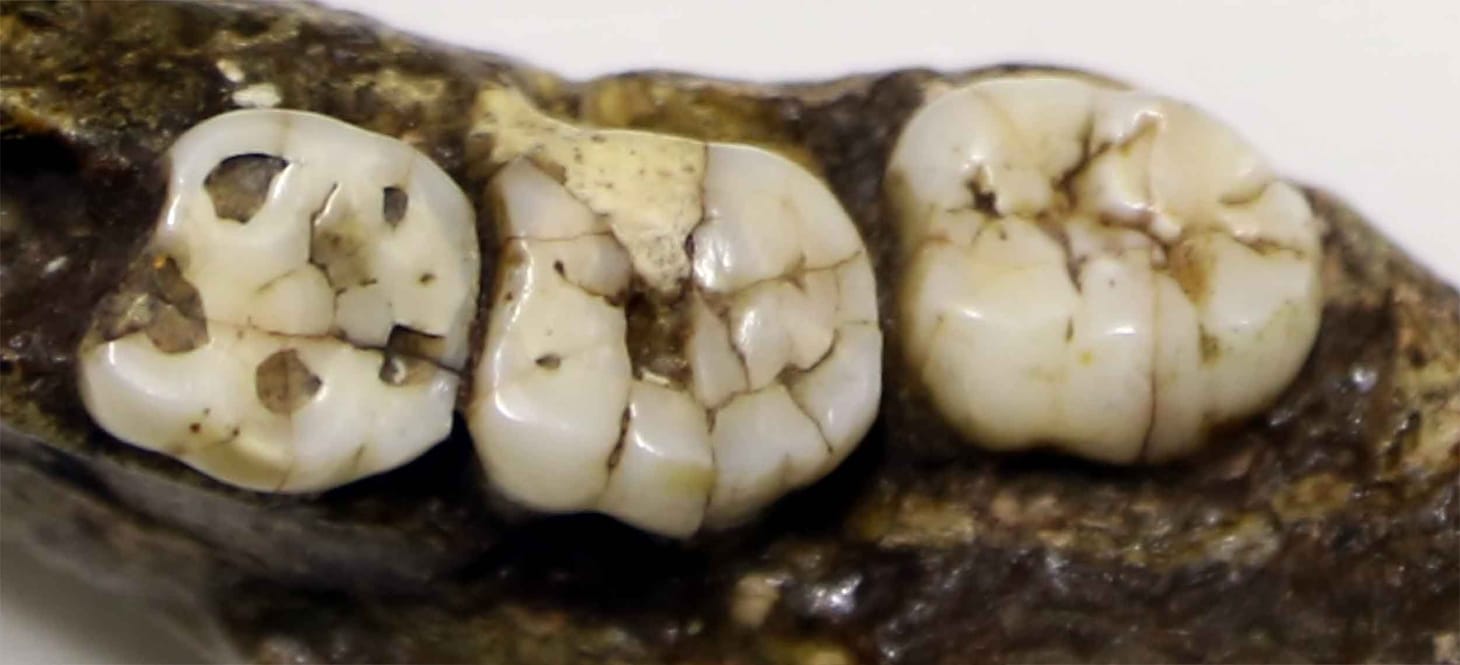
How many bathrooms have Neanderthals in the tile?
A Reddit poster finds an ancient jaw in his parents' new travertine. It may be more common than most people imagine.

Eclipses for the ancestors
Culture shapes our experience of these astronomical events, and would have done so for Neanderthals and other ancestral hominins.

A remembrance of Frans de Waal
Among many highlights of this primatologist's work, he maintained that humans are not unique or separated from other primates.

Vagrant birds and ancient human habitats
People killed the Carolina parakeet. An inquiry into their historic population range helps illustrate the challenges of understanding ancient human populations.

Guide to Paranthropus species
Long known as a group of human relatives with big teeth and jaws, these ancient species lived for at least two million years alongside our ancestors.

Secrets within the teeth of the first Homo fossils
New studies of the enamel-dentin junction show that early members of our genus may have been less distinctive than we think
Top 10 discoveries about ancient people from DNA in 2023
This year's highlights include ways of finding ancient relatives, how some phenotypes evolved in ancient people, and trace evidence from artifacts.
A visit to the Museo Nacional de Antropología, Mexico City
I took some time out from a research symposium and presentation to take photos of a marvelous collection of anthropological heritage.

All the hominins made tools
A study of associations between stone tool evidence and fossil hominin remains shows that a wide range of species made stone artifacts.
