Howler hybrid hunting
A montane hybrid zone between two species of Alouatta suggests an explanation rooted in cold tolerance.

Note: This post was written in 2007. It is an accurate description of the research published at that time and has been left with minor edits. However more recent research on these species of howler monkeys and research questions may be available.
Two species of howler monkeys, Alouatta palliata and Alouatta pigra, are sympatric within an overlapping part of both species' ranges in Guatemala. The two species are visibly different from each other: A. palliata, or mantled howlers, are mostly black with a lighter fringe of hairs on their flanks and back, while A. pigra, the Yucatán black howler, lacks this lighter coloration.
Today these species are rarely found in the same local area, with only small areas of overlap at the edges of their ranges. A. pigra lives in the Yucatan and adjacent areas of Mexico, Belize, and Guatemala. A. palliata lives in surrounding areas of Mexico, Honduras, and further south all the way to Ecuador. So, the range of A. pigra (black howlers) makes a sort of wedge in between two areas inhabited by A. palliata (mantled howlers). A. palliata is exceptionally cosmopolitan compared to the endemic A. pigra.
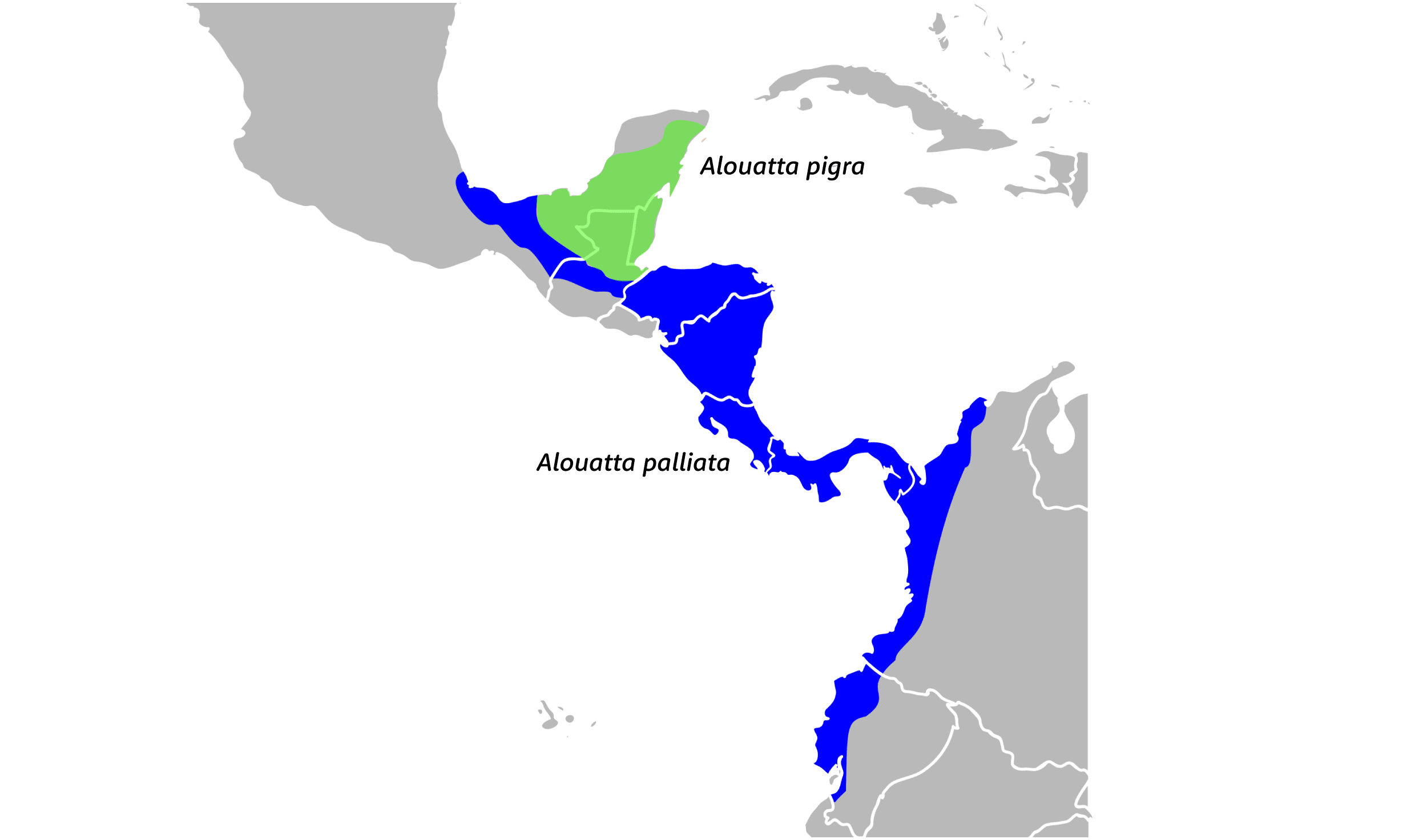
When the two species meet they do interbreed and give rise to visibly obvious hybrids. Liliana Cortés-Ortiz and colleagues sampled some DNA to figure out how permeable this hybrid zone really is. In previous genetic work, these authors have estimated the time of population divergence between A. palliata and A. pigra as 3 million years. When they looked at the genetics of hybrid individuals they found that the mixed individuals followed some classic predictions of population genetics theory.
These patterns imply that the direction of hybridization and subsequent backcrosses is strongly biased. Only crosses between A. pigra females or hybrid females carrying the mitochondrial haplotype of A. pigra and A. palliata males or hybrid males with the Sry gene of A. palliata occur and give rise to female offspring (Figure 3). However, no male hybrids with the Sry gene of A. palliata were observed and so, in accordance with Haldane's rule (HALDANE 1922), the data strongly suggest that the aforementioned crosses fail to produce viable males. Furthermore, on the basis of the low probability of detecting only the mtDNA haplotype of A. palliata in 12 adult hybrid individuals, A. palliata females and A. pigra males or hybrid males either mate infrequently or typically fail to produce viable offspring. Nonetheless, the genotypes of the hybrid infant and its suspected parents imply that this infant (S157, Table 2) was produced from a backcross between a male hybrid (S154, Table 2) and a female A. palliata. This demonstrates that such matings occur and that female offspring are produced. However, because no adult females were observed with the mitochondrial haplotype of A. palliata, we suspect that such crosses are uncommon or this infant is either infertile or will not survive to reproductive age.
I write a lot about the contributios of J. B. S. Haldane to the mathematical account of natural selection acting on genes. But another of his important—and probably more famous—contributions was the generalization about sex bias in interspecific hybridization.
How is it that the two species came to be split into different geographic ranges in this way? That is, why don't they overlap more than they do? Amrei Baumgarten (2006) noted that the two species have similar vegetational preferences, despite earlier reports that the more cosmopolitan A. palliata more readily invaded disturbed habitat such as banana and mango plantations. That suggests that it isn't dietary preferences, preferences for certain kinds of tree cover, or camouflage that is keeping them separate.
Instead, Baumgarten found that the primary difference between the two species is cold tolerance: A. pigra can and does live at higher altitudes than A. palliata, ranging high enough that it must tolerate freezing temperatures:
My study identified an important geographic barrier separating the two species: the highland massif of northern Central America, including Sierra Madre de Chiapas, Mexico, central highlands of Guatemala and Honduras (Fig. 3.1). The region is characterized by montane coniferous forests, subalpine forests and semiarid inland valleys, all unsuitable habitats for Alouatta. Therefore, the continuous highland belt in Mexico and Guatemala seems to be a barrier between the species and it defines the southern limit of the range of Alouatta pigra. A. palliata borders these mountains in Mexico at lower elevations in Tabasco, Veracruz and the Isthmus of Tehuantepec (Fig. 3.1). It probably bordered the same massif along its Pacific side as indicated by historical records in Mexico (Estrada and Coates-Estrada 1984), Guatemala (Handley 1950) and El Salvador (Daugherty 1972; USNM specimens # 282795, # 282850). (Baumgarten 2006:18).
To explain further, most of the Yucatan is relatively low in elevation. But the A. pigra–A. palliata barrier is primarily in the highlands to the south of the peninsula. No howlers live at the highest elevations, and the species' ranges are not contiguous in this area. Baumgarten (2006) proposes an interesting historical scenario to explain the current ranges. In his proposal, global cooling cycles that began around 2.7 million years ago may have retracted the howlers' rain forest habitat further south, leaving a relatively cold-adapted population ancestral to A. pigra in the Yucatan. Recurrent isolation and expansion of the two species may have occurred during intervening glacial and interglacial cycles.
Keeping this biogeographic hypothesis in mind, the comparative anatomy of the two species is very interesting. James Dale Smith (1970) reviewed the osteological differences between mantled and black howlers. Both species are considerably dimorphic in body size, and also have the distinctive dimorphism in the throat apparatus (including the hyoid bone) that distinguishes all howler monkeys. A. pigra is substantially larger than A. palliata—around 30 percent larger in mass (Ford and Davis 1992), with the difference more evident in males. This size difference is also reflected in the distinctive hyoid apparatus, which is much larger in the larger A. pigra males. The larger monkeys have more parallel tooth rows (contrasted with anteriorly converging rows in the smaller monkeys). Smith (1970) asserted that the most significant difference was in the maxillary molars. Large A. pigra skulls have molars that are roughly equal in size, while smaller A. palliata crania have molars that reduce in size from M1 to M3. The size differences are associated with a number of crown features, mainly related to simplification of the smaller M2 and M3 of the smaller monkeys.
Larger monkeys with larger, more complex molars, differences in throat anatomy, and greater cold tolerance, in contrast to a smaller, more cosmopolitan species, with the opportunity for gene flow during interglacials. They sound like Neanderhowlers.
Notes: This study was linked via Gene Expression.
References:
Baumgarten, A. (2006). Distribution and biogeography of Central American howling monkeys (Alouatta pigra and A. palliata). MA Thesis, Louisiana State University.
Cortés-Ortiz, L., Duda Jr, T. F., Canales-Espinosa, D., García-Orduña, F., Rodríguez-Luna, E., & Bermingham, E. (2007). Hybridization in large-bodied New World primates. Genetics, 176(4), 2421-2425. https://doi.org/10.1534/genetics.107.074278
Cortés-Ortiz, L., Bermingham, E., Rico, C., Rodrıguez-Luna, E., Sampaio, I., & Ruiz-Garcıa, M. (2003). Molecular systematics and biogeography of the Neotropical monkey genus, Alouatta. Molecular phylogenetics and evolution, 26(1), 64-81. https://doi.org/10.1016/S1055-7903(02)00308-1
Ford, S. M., & Davis, L. C. (1992). Systematics and body size: implications for feeding adaptations in New World monkeys. American Journal of Physical Anthropology, 88(4), 415-468. https://doi.org/10.1002/ajpa.1330880403
Smith, J. D. (1970). The systematic status of the black howler monkey, Alouatta pigra Lawrence. Journal of mammalogy, 51(2), 358-369.

John Hawks Newsletter
Join the newsletter to receive the latest updates in your inbox.


{kind=link}