The transition to Middle Stone Age from Acheulean did not make humans more deadly
Reading a meta-analysis of faunal data by Geoff Smith and coworkers that concludes that all Middle Pleistocene African peoples hunted the same prey animals.

The change in technology from Acheulean to Middle Stone Age in Africa was a major event in human prehistory. Or was it?
If there is one generalization that we can make from the African record, with all its imperfections, it is that large cutting tools were more common in assemblages from the earlier Middle Pleistocene than later. A second generalization is that Levallois-flaked points and evidence of hafting are more common in the later Middle Pleistocene than earlier. These may seem like a trade-off, and they may have been – although I’m not aware of anyone testing with numbers whether the two are causally connected instead of merely coincidental trends.
I think most anthropologists have assumed that technological change from Acheulean handaxes to Middle Stone Age hafted spears would make some difference in how ancient hominins hunted, scavenged, and ate animals.
The biggest-ever review of this question was published last year, with data drawn from sites across Africa from 800,000 to 130,000 years ago. The paper from Geoff Smith and coworkers, published in the Journal of Human Evolution, provides a useful and detailed review of faunal exploitation across African sites from the later Acheulean and earlier Middle Stone Age: “Subsistence strategies throughout the African Middle Pleistocene: Faunal evidence for behavioral change and continuity across the Earlier to Middle Stone Age transition”.
The review is a meta-analysis of published faunal data from site reports and articles. Looking at sites from Morocco across to Ethiopia and down through Kenya and Tanzania to South Africa, Smith and coworkers compiled faunal lists and other information from a varied array of more than 40 sites, some of which had faunal evidence from multiple time periods. They conclude that what seems like a big change in technology made no difference at all to which prey animals hominins hunted.
The currently available faunal data do not support a broadening of the hominin dietary niche during the Middle Pleistocene. While smaller-sized bovids, such as gazelles, grysbok, southern reedbuck, and springbok, are preserved throughout the Middle Pleistocene sample, these species never illustrate significant increases. Similarly, dangerous game (e.g., Cape buffalo and long-horned buffalo; see SOM Table S2) are recorded in small numbers throughout the faunal dataset. Increased proportions of dangerous game within MSA assemblages have been argued to reflect improvements in projectile technology, which provides the ability to hunt game at greater distances (Klein et al., 2007). While the proportion of these species increases from the early (0.03%) to late Middle Pleistocene (4%), they never predominate. Results from this metastudy suggest that such a change neither occurred on a broad scale during the Middle Pleistocene nor in parallel with the appearance of MSA technologies.
The data are pretty clear about two things. First, there is an enormous difference between open air and cave sites in faunal evidence.
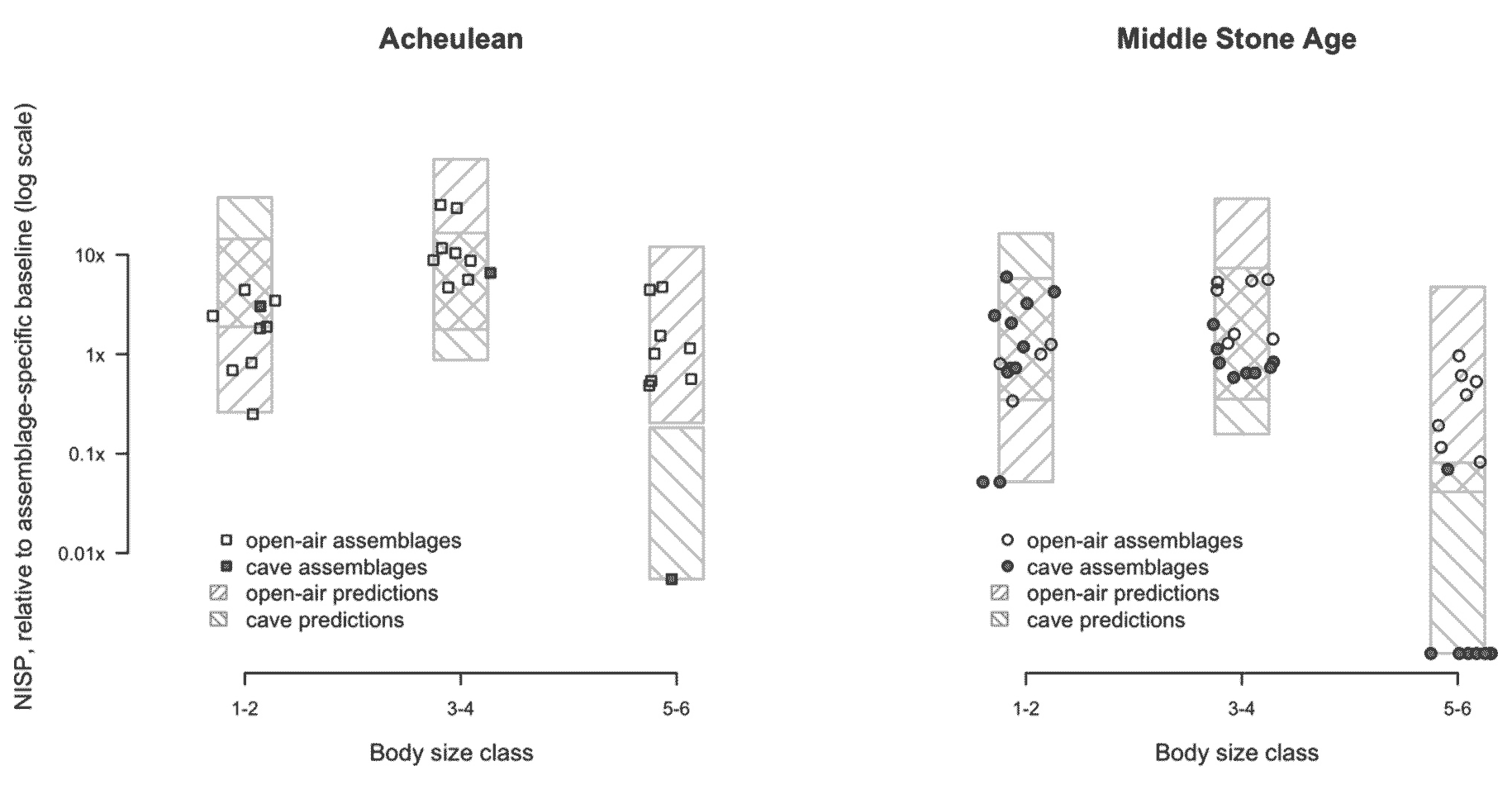
Hominins did not move the bones of elephants and hippos to caves, and they rarely moved large bovid remains. This is a known bias in zooarchaeology, but when you see a figure like this, it really reinforces just how much the hominin behavioral record is biased by the places that archaeologists prefer to dig. Some of the bias is that there are many cave sites in the MSA time period. Another bias is that archaeologists have been more likely to investigate open air sites that have obvious preservation of large mammal remains. A large fraction of hominin foraging behavior may be entirely missing from the record.
The other obvious feature of the results is that there really is no difference between Acheulean and MSA faunal size distributions other than the greater number of MSA sites that are caves.
A third important generalization from the data is that later Middle Pleistocene sites have more evidence for hominin processing of carcasses than earlier sites. The earlier sites have greater evidence for carnivore involvement, while later sites have more cutmarks and evidence for transport of selected body parts. However, that greater intensity of hominin involvement may reflect the bias toward cave sites in the later Middle Pleistocene.
There is too little data on mortality profiles across these sites for Smith and coworkers to do a comparison of selectivity of prime age adults in prey species. Still, there are many Oldowan-era sites that have a clear bias toward selection of prime age adults by hominins, at least for some prey size classes. It is not probably going to be very informative to look at more detailed aspects of prey selection without first knowing that the hominin contribution is much higher than carnivore contribution to faunal assemblages.
The meta-analysis approach obviously has limits. The quality of data is constrained by the data that has been reported. This varies greatly in depth and detail because some sites were excavated more than 50 years ago.
Also, the chronological information about sites is of varied quality. A good example is Kabwe, included here as a Sangoan/early MSA site. The recent paper by Rainer Grün and coworkers that provides some dates for the hominin material makes it clear that the contextual relationship of artifacts, faunal remains, and hominin fossils in the Broken Hill mine is basically unknown. It may be that the faunal material is of later Middle Pleistocene age, but it’s not obvious what (if any) hominin involvement may have been involved in accumulating animal bones. Kabwe is one of the rare sites that have both hominin fossil material, archaeological material, and fossil material. The unfortunate reality is that being from the same site does not provide evidence of association.
Other sites have similar difficulties. It may not seem objectionable acknowledge the uncertainty about geological age and simply lump sites as early or late. Still, the earlier open-air sites may be quite short duration accumulations that have very large uncertainty of geological age, while later cave sites may sample thousands of years with a more precise knowledge of age. Much rides on whether they are classified as “Acheulean” or “MSA”, yet that classification also depends in some cases on a small artifact assemblage. This kind of classification may serve the purpose of testing whether these coarse groupings have any correlation to faunal exploitation. But we cannot assume that such groupings are meaningful.
An enormous hole in this kind of project is the lack of knowledge of which (if any) hominins are responsible for the faunal remains. To their credit, Smith and coworkers are agnostic about which species of hominins are represented by the behavioral patterns across these many sites:
Currently, associated hominin fossils are limited and do not allow for direct correlations between various African Middle Pleistocene hominin species (e.g., Homo heidelbergensis, Homo rhodesiensis, Homo naledi and Homo sapiens) and a specific lithic technology or subsistence strategy. In general, this period is recognized by a mosaic of lithic entities and hominin species and the broad scale trends discussed here are complementary with more local and regional scale variation in terms of exact timings and causal mechanisms of behavioral change and continuity.
That’s an appropriately cautious statement. We do not know which hominin species made the artifact assemblages sampled at these sites. There were likely other species or divergent populations in addition to the ones listed. All of them were tool users and ate hunted animals.
With that caution in place, it remains interesting that there is no pattern over time across faunal assemblages in prey size selection. I think that’s striking evidence that earlier Middle Pleistocene hominins had achieved the ability to efficiently hunt prey species in the proportions that made ecological sense for them. The fact that later hominins selected the same prey sizes suggests that this aspect of hominin hunting was near equilibrium regardless of the precise toolkits they used.
If you ask me who was doing the hunting, I’d say it was every hominin that existed in Africa across this time period. I think it is likely that they all had similar niches with respect to carnivory.

John Hawks Newsletter
Join the newsletter to receive the latest updates in your inbox.