Two anthropologists float some curious notions about Homo naledi
I look at views expressed by Jeffrey Schwartz and Tim White about the anatomy of Homo naledi and its relationships with other hominins.
We’ve gotten lots of feedback on the new species Homo naledi. Most has been enormously positive, a little bit has been critical. In particular, a few scientists have come forward with criticism of the idea that H. naledi is really a new species. Fortunately I can address those criticisms easily by pointing to some easy-to-find answers.
Is H. naledi really two species?
On one side, Jeffrey Schwartz has argued that the H. naledi remains actually represent two or more different species. Newsweek gave him an op-ed platform to make his case, “Why the Homo Naledi Discovery May Not Be Quite What it Seems”.
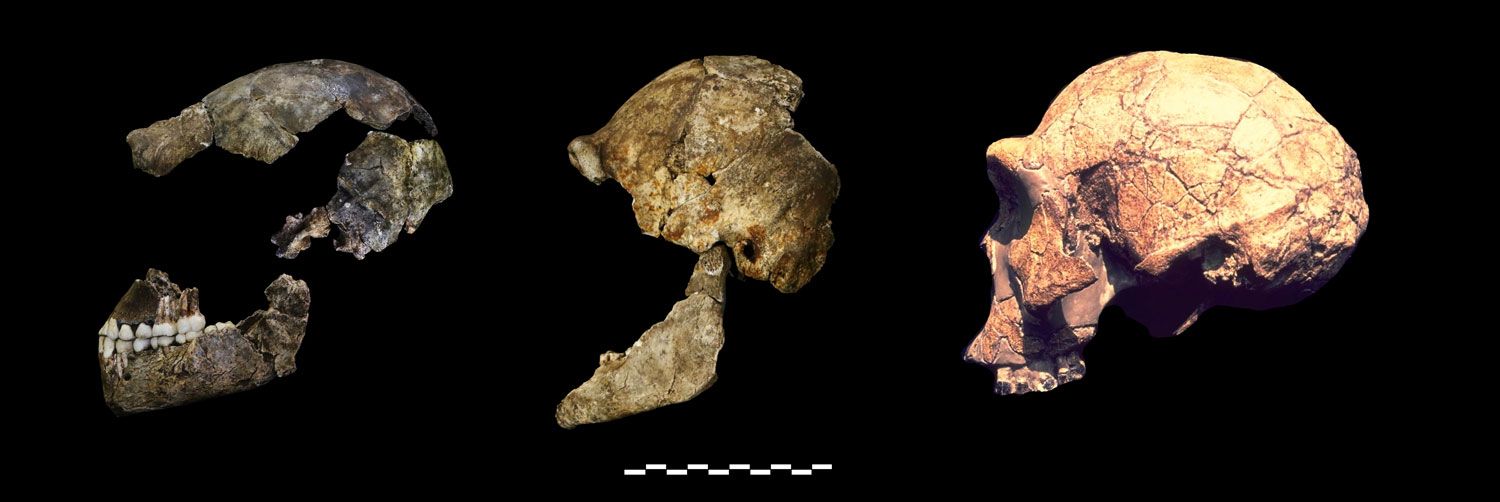
Viewed from the side, two partial skulls are long and low, with a long gently sloping forehead that flows smoothly into the brow – nothing like us, or most specimens regarded as Homo. A third partial skull is very short and rounded, with a high-rising forehead that is distinguished from a distinct, well-defined brow by a shallow gutter – not like the other skulls, and not like us or most specimens regarded as Homo. The femur has a small head (the ball end that fits in the hip socket) that is connected to the shaft of the bone by a long neck, and, below the neck, is a "bump" of bone that points backward. These features are seen in every australopith femur. In us, and all other living primates, the head of the femur is large and the neck short, and the "bump" points inward. Further, the teeth are very similar to those from a nearby fossil site that has yielded various kinds of australopith. Even at this stage of their being publicized, the "Homo naledi" specimens reflect even greater diversity in the human fossil record than their discoverers will admit.
Homo naledi has a mosaic of features that include some that compare most closely to more primitive australopiths, and others that compare more closely to Homo. How do we know that this is one species rather than a jumble of species mixed together? Simple: every feature that is repeated in the sample is nearly identical in all individuals that preserve it. It would be very strange to have a mix of different species where all seven proximal femora come from one species, while all of a dozen lower third premolars come from a different species.
Schwartz is also totally wrong about the crania: All the frontal bones in the collection have a similar morphology with a thin supraorbital torus and slight but distinct supratoral sulcus. The variation within the collection is not high, it is extraordinarily low—in fact, the tooth size varies less than half as much as is usual in most human populations. Schwartz seems to imply in his argument that some of the Swartkrans teeth attributed to Homo are similar to H. naledi. There are some superficial similarities, but in fact we cannot find a good comparable example within the Swartkrans collection, particularly for the distinctive premolar crown and root morphology, the lower canine crown morphology, and the small molar sizes of H. naledi.
All of that is in the paper, which is open access for anyone to read anywhere in the world: “Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa”. It is not really a mystery why someone like Schwartz would come forward in such a public way with criticisms that are easily answered by the open access paper—after all, it’s not every day that an anthropologist is asked to write a full-page piece for Newsweek.
The “primitive small Homo erectus” idea
Another criticism of Homo naledi has come from a prominent paleoanthropologist and member of the National Academy of Sciences, Tim White. White has been publicizing his theory about Homo naledi in many press interviews (e.g., “Some bones to pick”):
White: Dating is irrelevant; these are a small, primitive H. erectus, whatever the date turns out to be. This is because they are not biologically different, in any significant way, from already known H. erectus found in places like Swartkrans (800m away), eastern Africa, or the Georgian republic. Of the 80+ traits listed in the e-LIFE supplemental material, only a small fraction of them are even claimed to differentiate these fossils from earlier described H. erectus, and that fraction of characters is known to vary among members of the same species (even population) of both H. erectus and H. sapiens. In other words, the newly described "species" is an example of artificial species inflation in palaeoanthropology.
Of course, Homo erectus is a very well-known species, and we compared the Dinaledi fossils to every specimen available to us. Many of our team have studied the originals of most Homo erectus remains around the world, and I have personally examined the key cranial and postcranial specimens from Dmanisi. So we examined this question in great detail as we studied the Homo naledi material.
How are we to know that Homo naledi is not the same as a primitive, small Homo erectus? Well, for one thing, at least two H. naledi individuals have endocranial volumes around 460 cc, much smaller than any H. erectus cranium ever found. There is barely any overlap between the larger individuals and H. erectus, with only a single H. erectus specimen coming close to the H. naledi range of variation in volume.
White doesn’t specify how many features would be sufficient in his view to define a species. It should be obvious that if we list 80 cranial and dental traits that are informative among all hominins, that most species will differ in only a relatively small fraction of those. Nevertheless our open access paper lists many clear differences between H. naledi and H. erectus, including aspects of premolar crown and root morphology, the crown morphology of the molars (simplified in H. naledi, invariably crenulated and complex in H. erectus), vault shape (H. naledi does not have the elongated, low cranium of H. erectus) and mandibular shape—all listed on page 10 of our open access paper. Even setting aside the postcranial skeleton and the very small endocranial volume, these show H. naledi to be distinct from H. erectus.
When we look at the postcranial skeleton, there is simply no way that H. naledi could be confused for H. erectus. H. naledi has a long, anteroposteriorally flattened and anteverted femur neck, which looks very different from African and Dmanisi femora attributed to H. erectus. The H. naledi tibia is exceptionally mediolaterally thin and long, with a rounded anterior border and tubercle for the pes anserinus tendon, all traits that we could not find in known tibiae attributed to H. erectus including Dmanisi. The H. naledi scapula has a superiorly oriented glenoid, very different from the Dmanisi scapula specimen or the Nariokotome H. erectus skeleton. The vertebrae of H. naledi do not match in proportions or morphology the comparable examples from Nariokotome or Dmanisi, and the pelvis of H. naledi exhibits a short, flared ilium unlike those known for H. erectus, including the Gona pelvic specimen.
It’s just a poor match to H. erectus, so that the only way to make the H. naledi fossils fit within Homo erectus is to stretch that species beyond any other ever defined in the human lineage. There are clearly some similarities, which to us indicate something about the evolutionary relationships of H. naledi and H. erectus—but we are hesitant to go so far as to posit a unique relationship of these two species because many of their similarities also overlap with species such as H. habilis, H. rudolfensis, and even Australopithecus sediba. Figuring out those relationships will take additional research and analysis.
What a species diagnosis does
The purpose of a species diagnosis is to enable other researchers to distinguish the new species from other species they already know. The diagnosis is a taxonomic act, creating a name that can be recognized in the future by agreed-upon rules of nomenclature. The species diagnosis provides a basis for later descriptive work, which can go into much more detail about the morphology of the fossils and include comparisons with all specimens of other species rather than merely differential traits.
It has been tradition in paleoanthropology for the first diagnosis of a new hominin species to be very brief. For example, the diagnosis and description of Australopithecus ramidus (White et al. 1994) included dental, cranial and postcranial material, and was only 6 pages long. The diagnosis and description of Australopithecus garhi (Asfaw et al. 1999) was also 6 pages. The diagnosis and description of the new species Australopithecus deyiremeda (Haile-Selassie et al. 2015) is 5 pages.
Of course there is a problem with such short descriptions. How were we really to know that Australopithecus ramidus was different from Australopithecus afarensis? More to the point, a year later, when White and colleagues published an erratum to their article that named a new genus for the material, which they called Ardipithecus, how were we to know that was justifiable based on evidence? Well, in that case we waited 15 years for the publication of additional descriptive material. The description of Au. garhi is still, 16 years later, the only scientific document on those fossil remains. The description of Au. deyiremeda this spring (Haile-Selassie et al. 2015) was met with immediate criticism by Tim White, who seemingly has no time for species that he himself has not named.
Some people have become jaded about the entire idea of hominin species, cynically concluding that every new species name is just an exercise in glorifying the discoverer, or is stoked by nationalist pride. In my experience, that’s rarely true. Paleoanthropologists are careful scientists who want their work to stand the test of public attention. But the field’s convention of brief species diagnosis followed by long delays for more information has led to confusion and criticism.
We hope that a more open approach will lead to more productive conversations about hominin species.
With 35 text pages, not counting supplementary data, the species description of Homo naledi is around 6 times longer than has been typical in paleoanthropology. We worked to give a much broader perspective on the anatomy across the skeleton than is typical of species descriptions in our field, and we have described the differences between H. naledi and other species with much greater specificity than has been done before.
There is much work to do in describing the Homo naledi collection, and we have several publications forthcoming and in review that give more descriptive detail about the H. naledi fossils. This is just the beginning of a conversation about the form and place of this species in our evolutionary history. But we think that the data we have provided in this initial publication will allow people to know with some confidence that H. naledi is really quite different from other previously-known species.
Download them yourself!
Of course, no one has to trust what we have written about the fossils, because we have made the 3D shape files for many of them available open access through MorphoSource. Anyone can sign up for a free login and download the shape files, even print them out. It’s been exciting to see anthropology departments and seminars printing the fossils and discussing them all week long!
Some senior paleoanthropologists have unfortunately been accustomed to secretive practices. They may think that people will trust their authoritative pronouncements about fossil remains because no one will ever see the data. That’s an unscientific approach, and it leads to bad practices.
What we’re seeing now from senior scientists like Tim White and Jeffrey Schwartz is an unfortunate pattern. These scientists are used to being able to immediately talk to their cronies in the press with a knee-jerk reaction to new fossil publications, secure in the knowledge that the authors won’t release the data to contradict them. Even White, who has famously written (White 2000) that no one should publish on a fossil without seeing the original, has twice this year chased attention for his own fringe views about new species.
I hope that he’ll print out the fossils and take a careful look side-by-side with his own data. Maybe someday he’ll share his data with other scientists and the world so that we can have a real conversation.
References
Asfaw, Berhane, et al. "Australopithecus garhi: a new species of early hominid from Ethiopia." Science 284.5414 (1999): 629-635.
Berger, Lee R., et al. "Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa." eLife 4 (2015): e09560.
Haile-Selassie, Yohannes, et al. "New species from Ethiopia further expands Middle Pliocene hominin diversity." Nature 521.7553 (2015): 483-488.
White, Tim D., Gen Suwa, and Berhane Asfaw. "Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia." Nature 371 (1994): 306-312.
White, Tim D., Gen Suwa, and Berhane Asfaw. "Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia." Nature 375.6526 (1995): 88.
White, Tim D. News and Views: A View on the Science: Physical Anthropology at the Millennium. American Journal of Physical Anthropology 113 (2000):287–292.

John Hawks Newsletter
Join the newsletter to receive the latest updates in your inbox.


